The continued increase in incidence of some hormone-related cancers worldwide is of great concern. Although estrogen-like substances in the environment were blamed for this increase, the possible role of endogenous estrogens from food has not been widely discussed. We are particularly concerned about cows� milk, which contains a considerable quantity of estrogens. The milk that we are now consuming is quite different from that consumed 100 years ago. Unlike their pasture-fed counterparts of 100 years ago, modern dairy cows are usually pregnant and continue to lactate during the latter half of pregnancy, when the concentration of estrogens in blood, and hence in milk, increases. The correlation of incidence and mortality rates with environmental variables in worldwide countries provides useful clues to the etiology of cancer. In this study, we correlated incidence rates for breast, ovarian, and corpus uteri cancers (1993-97 from Cancer Incidence in Five Continents) with food intake (1961-97 from FAOSTAT) in 40 countries. Meat was most closely correlated with the breast cancer incidence (r=0.827), followed by milk (0.817) and cheese (0.751). While stepwise multiple-regression analysis (SMRA) identified meat as the factor contributing most greatly to the incidence of breast cancer ([R]=0.862), the dietary factor identified as the most significant contributor to the mortality was milk plus cheese ([R]=0.814). Milk was most closely correlated with the incidence of ovarian cancer (r=0.779), followed by animal fats (0.717) and cheese (0.697). SMRA revealed that milk plus cheese make the greatest contribution to the incidence of ovarian cancer ([R]=0.767). Milk was most closely correlated with corpus uteri cancer (r=0.814), followed by cheese (0.787). SMRA revealed that milk plus cheese make the most significant contribution to the incidence of corpus uteri cancer ([R]=0.861). In conclusion, increased consumption of animal-derived food may have adverse effects on the development of hormone-dependent cancers. Among dietary risk factors, we are most concerned with milk and dairy products, because the milk we drink today is produced from pregnant cows, in which estrogen and progesterone levels are markedly elevated.
Introduction
Breast cancer is one of the most frequent cancers among women (1). The incidence of breast cancer has been increasing in both developed and developing countries (2). The fact that people from low-risk countries increase their risk on immigrating to higher-risk countries suggest that some modifiable lifestyle and/or environmental factors are responsible for the development of breast cancer (3). It has been postulated that most cancer-afflicted women develop subclinical breast cancer at an early age and that diet plays an important role in the progression from such lesions to clinical disease (4). In addition, the variation in the breast cancer incidence rate worldwide suggests that lifestyle factors, especially diet, influence breast cancer risk.
Epithelial ovarian cancer kills more women each year than all other gynecological cancers combined (5). The incidence of ovarian cancer also varies widely among countries. In general, incidence rates are higher in developed than in less developed countries. Rates are higher among younger Chinese and Japanese women born in the United States than among their Asian-born counterparts (6). Such immigration studies suggest that lifestyle and environmental factors play a major role in the etiology of this malignancy.
Endometrial cancer is the sixth most common cancer worldwide, accounting for ~2% of all cancers in women (7). As with breast and ovarian cancers, there is a wide difference in the incidence rate of corpus uteri cancer across countries. The rate amongst U.S. Japanese is several-fold higher than that for Japanese living in their native country (3). This finding suggests that lifestyle and environmental factors, rather than hereditary influence, affect the incidence of this cancer.
The common feature of breast, ovarian, and corpus uteri cancers is that they are comparatively rare in women under 40 years of age. The risk increases sharply after around age 40 and peaks between the ages of 65 and 79 (8). The three malignancies share common risk factors, including obesity, menarche at young age, late menopause, and nulliparity, in addition to an increased risk inherent to and dependent upon family history (5). In addition, the risk of these three cancers changes within a few years of shifts in exposure to sex hormones, and some of the changes in risk persist for many years, indicating that hormones can affect both early and late stages of carcinogenesis. Sex hormones may play a major role in their etiology, likely by controlling the rate of cell division, cell differentiation, and number of susceptible cells (9).
According to Armstrong and Doll (10), the correlation of incidence and mortality rate with the prevalence of environmental agents in various geographical areas provides useful clues to the etiology of cancer. In this study, we used incidence and mortality data for the three most prevalent female cancers �breast, ovarian, and corpus uteri cancers from 40 countries� and correlated the rates with dietary variables.
Materials and Methods
Cancer
incidence
Cancer Incidence in Five Continents edited by Parkin et
al. (11) provided us with comparable data on the incidence of cancer between
1993 and 1997 in different geographical locations (200 populations in
50 countries). In this study, we used data from 40 of these countries,
for which both cancer incidence rates and food consumption data from FAOSTAT
Database Collections (12) were available. The age-specific incidence rates
were standardized to the world population and the mean value of the age-adjusted
incidence rates were calculated.
Several countries have more than one cancer registry. If more than one registry was available for a country, those that stated �The editors were unable to verify these data� were excluded from this study. The remaining age-specific incidence rates in the same country were standardized to the world population. The mean age-adjusted incidence rate was used as a representative rate for Australia, Brazil, China, France, Germany, India, Italy, Japan, Poland, Spain, Switzerland, Thailand, and the United Kingdom.
In some countries, cancer registries provided data for different ethnic groups. In this study, the rates for Africans were used for Zimbabwe; those for all Jews were used for Israel; those for Kuwaitis were used for Kuwait; and those for non-Maori were used for New Zealand.
In several
countries, only one cancer registry reported data to the IARC. The incidence
rate from the registry was then assumed to represent the incidence for
the whole country (Algeria, Argentina, Austria, Columbia, Costa Rica,
Ecuador, Ireland, Korea, Mali, Malta, Philippines, Uganda, Uruguay, and
Viet Nam).
Canada, Denmark, Finland, Hong Kong, Iceland, The Netherlands, Norway,
and Sweden collected data for their cancer registries on a national basis.
The incidence rates for these countries were used as provided.
Many cancer registries operate in the United States. Of these, the registry
for Caucasians produced by the Surveillance, Epidemiology, and End Results
(SEER) program was used as representative for the United States.
The age-specific mortality rates of cancers that are listed in GLOBOCAN
2000 (13) were used in this study. The countries selected were the same
as those used for the incidence.
Food
consumption
The consumption data (Mt/1000 capita/year) for various food items
from 1961 to 1997 available from the FAOSTAT Database Collections (12)
were used. The food items used for this study were animal fats, meat (beef,
pork, poultry, mutton, and goat meat), eggs, butter, milk (excluding butter),
whole milk, cereals (excluding wine), pulses, beans, soy beans, peas,
fruits (excluding wine), vegetables, coffee, tea, and alcoholic beverages.
The consumption of each food or drink was converted from Mt/1000 capita/year
to g/capita/day.
Statistical analysis
All of the data were analyzed by a statistical software (SAS Institute,
Cary, NC). The 0.05 level of probability was used as the criterion for
significance. Simple correlation coefficients (r) were calculated to examine
the association between the incidence or mortality rates of breast, ovarian,
and corpus uteri cancers and the consumption of each food item. Consumption
of several food items was closely interrelated (collinearity, r > 0.8).
For example, the r value calculated for the correlation between the amount
of milk and cheese consumed in 1961-97 was 0.812. Hence, consumption of
milk and cheese in the same period was grouped as �milk + cheese.� The
contributions of these grouped food items to incidence or mortality were
evaluated by stepwise multiple-regression analyses. The following eleven
food items were selected for the analysis (independent variables): �animal
fats + butter,� �milk + cheese,� eggs, meat, cereals, pulses, fruits,
vegetables, vegetable oils, coffee, and alcohol.
Results
Incidence
of breast cancer and food intake
The age-adjusted incidence rates per 100,000 for breast cancer differed
markedly from one country to another. For example, Uruguay had the highest
rate at 114.9, followed by the United States (92.1) and Israel (87.1)
(Figs. 1 and 2). The lowest rate was for Korea (12.7), followed
by Thailand (16.1) and Mali (20.0).
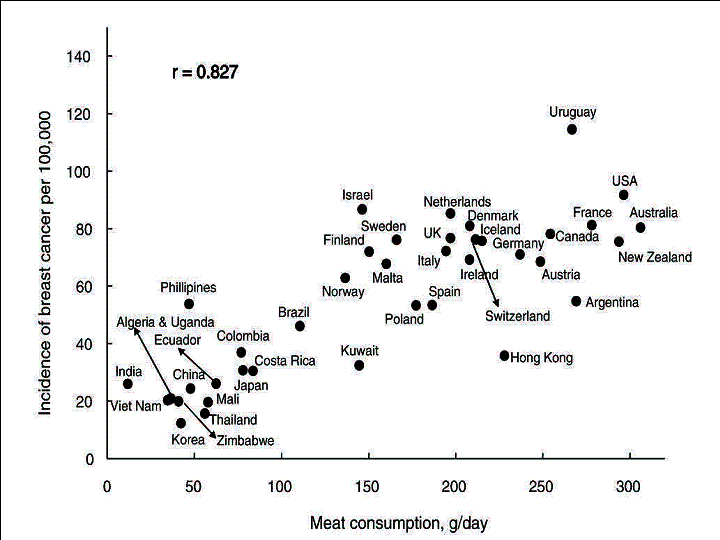
Fig.
1. Correlation between the age-adjusted incidence of breast cancer and
per capita meat consumption.
�@
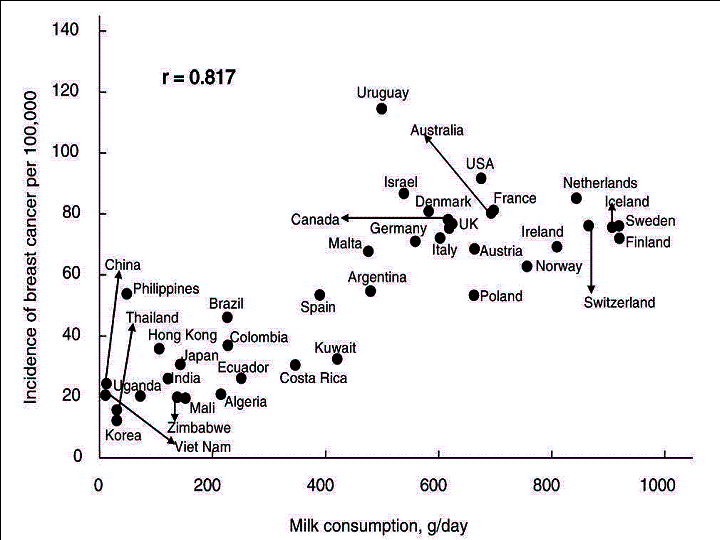
Fig.
2. Correlation between the age-adjusted incidence of breast cancer and
per capita milk consumption.

The simple correlation coefficients between the incidence rate of breast cancer and food intake in 40 countries are shown in Table 1. Of the food items examined, meat was the most closely correlated with incidence of this cancer incidence (r=0.827) (Fig.1), followed by milk (0.817) (Fig. 2) and cheese (0.751). By contrast, cereals (-0.467) and pulses (-0.438) were negatively correlated with the incidence of breast cancer.
Stepwise multiple-regression analysis revealed that meat (1961-97) contributed most to the incidence of breast cancer in 1993-97 (standardized regression coefficient [R]=0.862) (Table 2).

The correlation coefficient between the age-adjusted incidence rate and age-adjusted mortality rate of breast cancer in the 40 countries examined was r=0.872. The age-adjusted mortality rates of breast cancer had the highest correlation with cheese consumption (r=0.725), followed by fats (0.670) and butter (0.595) (Table 3).
Incidence
of ovarian cancer and food intake
Iceland had the highest ovarian cancer incidence at 16.2 per 100,000,
followed by Sweden (15.2) and the United Kingdom (13.7) (Fig. 3).
The lowest rate per 100,000 was for Korea (1.6), followed by Mali (2.1),
China (4.0), and Brazil (4.0).
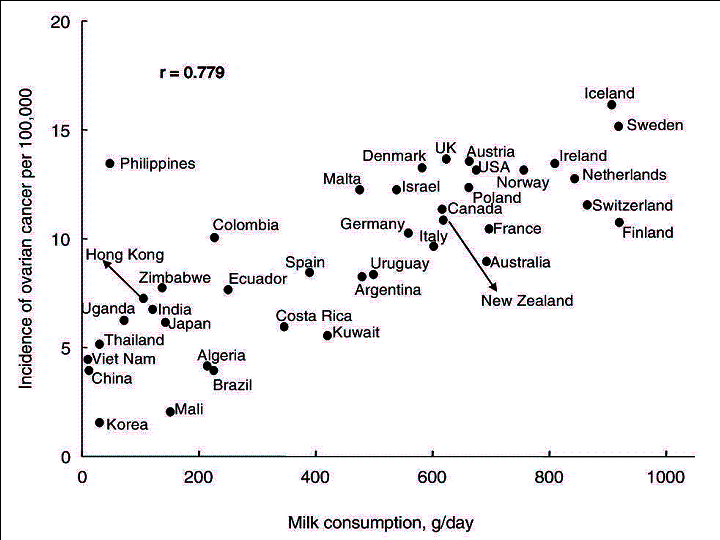
Fig.
3. Correlation between the age-adjusted incidence of ovarian cancer and
per capita milk consumption.
The simple correlation coefficient, r, showed the greatest correlation between milk and ovarian cancer (r=0.779), followed by animal fats (0.717) and cheese (0.697) (Table 1). By contrast, cereals (r=-0.520) and pulses (-0.465) were negatively correlated with the incidence of ovarian cancer.
The results of stepwise multiple-regression analysis are presented in Table 2. The analysis identified �milk + cheese� as the factor contributing most to the incidence of ovarian cancer (standardized regression coefficient [R]=0.767), while it identified cereals as a factor contributing to a decreased incidence.
The correlation coefficient between mortality and incidence of ovarian cancer was as high as r=0.808. The food most closely correlated with mortality due to ovarian cancer was fats (r=0.818), followed by milk (0.790) and cheese (0.731) (Table 3). Cereals, by contrast, were negatively correlated with the mortality (-0.391).
Incidence
corpus uteri cancer and food intake
The age-adjusted incidence rate (per 100,000) of corpus uteri cancer was
highest in the United States (18.4), followed by Malta (16.3) and Canada
(14.4). Viet Nam had the lowest incidence rate at 1.6, followed by Mali
(1.7) and Korea (1.9) (Fig. 4).
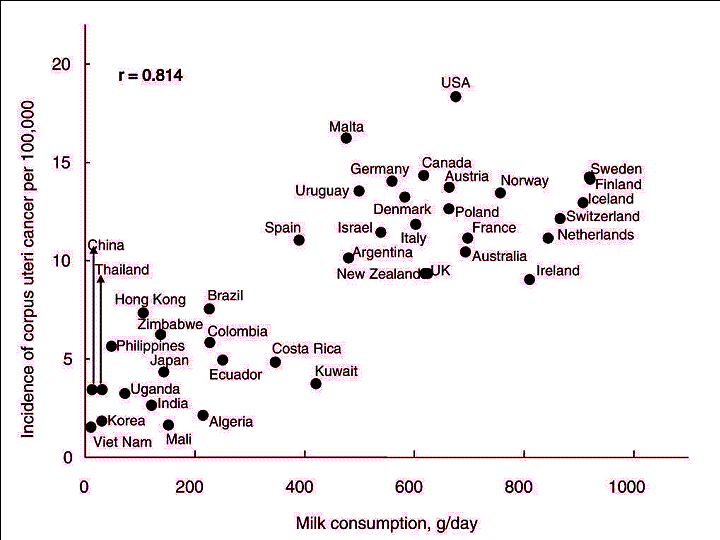
Fig.
4. Correlation between the age-adjusted incidence of corpus uteri cancer
and per capita milk consumption
Milk was most closely correlated with the incidence of corpus uteri cancer (r=0.814), followed by cheese (0.787) and meat (0.782) (Table 1). Conversely, pulses were negatively correlated with the incidence of this cancer (-0.437).
Stepwise multiple-regression-analysis identified �milk + cheese� as the factor contributing most to an increased incidence of corpus uteri cancer (standardized regression coefficient [R]=0.861) (Table 2).
The mortality rates of corpus uteri cancer were weakly correlated with its incidence rate (r=0.251). The highest age-adjusted mortality rate was 4.4 for Uruguay, and Mali had the lowest rate of 0.24. As with the incidence data, the age-adjusted mortality rates of this cancer had the highest correlation with milk consumption (r=0.545), followed by cheese (0.406) and animal fats (0.403) (Table 3).
Discussion
Epidemiological studies that correlate cancer incidence and mortality rates with the dietary practices in various geographical areas have certain shortcomings. The rates of incidence and mortality of any cancer are affected by regional differences in diagnosis, registration and certification, and the fatality rate of the cancer. The cancer incidence and mortality data from developing countries are clearly less complete than those from developed countries because of problems with underdiagnosis and undercertification due to the local medical and economic background, together with problems enumerating the population. In addition, the consumption data for each food item in a country are not based on actual nutrition surveys, but are roughly estimated using the following equation: food supply per capita = (production + import + stock changes - export - feed - seed - processing - waste - other uses)/population. Hence, it is difficult to determine the extent to which the available food data translates into daily per capita consumption. Nevertheless, the overall relationships are convincing, plausible, and at least serve as guides for further epidemiological and experimental studies (10,14).
Breast
cancer
Early diagnosis (mammography, fine-needle biopsy), treatment (lumpectomy,
radiotherapy, chemotherapy) and increasing public awareness of breast
cancer may have contributed to its relatively stable mortality rates despite
its increasing incidence. Starting in the 1990s, breast cancer mortality
began to decline in the United States and in some European countries (8).
Since the 1950s, however, both incidence and mortality rates have been
increasing in a number of other countries, including low-risk countries
such as China (15) and Japan (16). Lifestyle changes in these countries,
such as diet, likely account for much of this increase.
In this study, the food that was most closely correlated with the incidence of breast cancer was meat, followed by milk and cheese. Many other epidemiological studies have found positive correlations between breast cancer risk and the consumption of meat, milk, and dairy products (14,17-21). According to La Vecchia and Pampallona (18), who found a significantly positive correlation between breast cancer mortality and meat and milk consumption, milk and cheese were the only dietary variables to remain significantly positive after the correlation was adjusted for the women�s age at the birth of their first child and economic variables.
Since dietary fat has long been considered a major risk factor for breast cancer, special attention has been given to the high fat content of milk (17,19,21-23). However, milk fat cannot account for all of the breast cancer risk related to milk consumption. In fact, consumption of whole milk has declined steadily since the 1950s, and been replaced by fat-reduced milk (24), yet the incidence of breast cancer has increased worldwide over the last 50 years (25).
Estrogens and progesterone are breast cancer promoters because of their physiological roles stimulating the mammary gland (26). The major sources of animal-derived estrogens in the human diet are milk and dairy products, which account for 60-80% of the estrogens consumed (27). Most milk for human consumption is obtained from heifers in the latter half of pregnancy, when the estrogen levels in cows are markedly elevated (28).
Although
the oral bioactivity of free estradiol and estrone may be relatively low,
estrone sulfate, which is a major estrogen in milk, has high oral bioactivity.
Once inside the body it can be readily converted to estrone and estradiol.
Postmenopausally, estrone is the source of most circulating estradiol,
and estrone sulfate is the most abundant circulating estrogen (29). In
addition to the long plasma half-life of estrone sulfate, it is absorbed
in the intestinal mucosa either virtually unchanged (30,31) or by enterohepatic
circulation. Remesar et al. (32) calculated estrone intake in a standard
human diet and found that 46.6% of estrone came from dairy products.
Recently, we confirmed that commercially available low-fat milk promotes
the development of 7,12-dimethylbenz[a]anthracene (DMBA)-induced mammary
tumors in rats (33). In order to specify the role of estrogens on the
development of mammary tumors, an aqueous estrone sulfate solution was
used as a positive control in this study. The degree of tumor promotion
by cows� milk was almost comparable to that of 100 ng estrone sulfate
per milliliter.
Some human studies have also demonstrated that milk consumption increases the circulating estrogen level. South African black males who had switched from a vegetarian diet to a Western diet with milk, butter, and meat showed an increase in circulating estrogen levels (34). The relationship between milk consumption and plasma estrogen concentrations is also supported by the fact that Asian women, whose consumption of milk and dairy products is low, have lower plasma estrogen concentrations than those of Caucasian women, whose dairy product consumption is high (35).
In addition to estrogens, milk also contains insulin-like growth factor I (IGF-I) in nanomolar concentrations (36). IGF-I plays a role in the growth and incidence of neoplastic pathology (37) by regulating the growth of cancer cells and it stimulates the proliferation of human breast cancer cell line MCF-7 (38). Further, estrogen and IGF-I have mutually favorable actions on breast cancer cell proliferation. In a human study, plasma IGF-I concentration increased by 10% when healthy subjects consumed cows� milk (39). In fact, the consumption of commercial milk pasteurized by heat at 125-130 �C was confirmed to increase the plasma level of IGF-1 in rats (33).
Obesity has been linked with breast, corpus uteri and ovarian cancers, probably via its effect on circulating estrogens (40,41). This finding is biologically plausible, since a greater amount of fat results in greater conversion of androstenedione to estrone by aromatase and lower levels of sex hormone-binding-globulin (SHBG), which leads to higher levels of bioavailable estrone and estradiol (42). We should pay particular attention to the sex hormone in modern cows� milk, because epidemiological studies of large scale have repeatedly confirmed the increased incidence of breast cancer in postmenopausal women who are currently undergoing hormone replacement therapy (43,44).
Ovarian
cancer
The strong correlation between ovarian cancer and per capita milk consumption
led to the hypothesis that consumption of lactose is a risk factor for
ovarian cancer (8). In the Iowa Women�s Health Study, a prospective study
of 41,836 women, high lactose consumption was associated with an increased
risk of ovarian cancer (r=1.60) (45). Cramer (46) proposed that galactose,
a component sugar of lactose, is a risk factor for ovarian cancer, as
galactose is toxic to oocytes. Hypogonadism or ovarian failure in response
to gonadotropin stimulation has been found in women who are deficient
in the enzyme galactose-1-phosphate uridyltransferase (GALT) (47). In
one case-control study, GALT activity was significantly lower in cases
of ovarian cancer (48). Nevertheless, most case-control studies that have
examined the risk of ovarian cancer in relation to lactose intake have
not reported a substantial association between the two (49,50). A recent
prospective cohort study in Sweden (51) indicated that high intakes of
lactose and dairy products, particularly milk, are associated with an
increased risk of serious ovarian cancer but not of other subtypes of
ovarian cancer.
Several research groups have reported associations between ovarian cancer and the intake of specific milk products, such as butter (52,53), whole milk (53-55), skim milk (45), yogurt (48-55), cottage cheese (48), and ice cream (53). One study found a three-fold greater risk of ovarian cancer among women who drank at least one glass of whole milk per day compared with those who did not (54). Risk was also elevated for non-fat milk, eggs and cheese, and dairy products in general (45).
In our study, the stepwise multiple-regression analysis revealed that �milk + cheese� contributed the most to the incidence of ovarian cancer. Our results are consistent with previous ecological studies that found an association between ovarian cancer and milk consumption (14). The etiology of ovarian cancer is still poorly understood, although it is clear that hormonal factors are involved. Higher IGF-I levels and reduced IGF-I binding protein levels have also been found in patients suffering from ovarian cancer (56,57). Epidemiological observations and experimental data indicate that estrogens have an adverse effect on the ovarian epithelium (7). It is not unreasonable to presume that estrogens and growth factors in milk and dairy products are associated with the development of ovarian cancer by changing the hormonal environment in the ovaries.
Corpus
uteri cancer
Cancer of the corpus uteri is a typical model of hormone-related carcinogenesis.
Indeed, the effect of estrogens is so direct and strong that the widespread
use of hormone therapy increased the incidence of this cancer in the United
States to almost epidemic proportions (8). Most of the risk factors for
corpus uteri cancer can be explained within the framework of the unopposed
estrogen hypothesis, which holds that exposure to estrogens leads to increased
mitotic activity of endometrial cells, an increased number of DNA replication
errors, and somatic mutations resulting in a malignant phenotype (58,59).
In our study, the incidence of corpus uteri cancer was most closely correlated with the consumption of milk and cheese. As discussed in the section on breast cancer, milk and dairy products contain considerable amounts of estrogens and growth factors. Estrogens induce various tumors in laboratory animals and have been recognized as carcinogens in humans, raising the risk for uterine cancer, in addition to breast and ovarian cancers (60). Several trials have established a link between endometrium cancer and the IGF system (61-63), including the stimulating effect of IGF-I in vitro on endometrial cancer cells, and increased circulating levels of serum IGF-I and decreased insulin-like growth-factor-binding protein (IGFBP-1) in women with corpus uteri cancer.
In conclusion, we hypothesize that female sex hormones in milk and dairy products have a common effect on the development of breast, ovarian and corpus uteri cancers. Further epidemiological and mechanistic studies are needed to verify this hypothesis.
References
1.
Parkin DM. The global burden of cancer. Semin Cancer Biol 1998;8:219-35.
2. Coleman MP, Esteve J, Damiecki P, Arslan A, Renard H. Trends
in cancer incidence and mortality. IARC Sci Publ 1993;121, Lyon, IARC.
3. Ziegler RG, Hoover RN, Pike MC, et al. Migration patterns and
breast cancer risk in Asian-American women. J Natl Cancer Inst 1993;85:1819-27.
4. Woutersen RA, Appel MJ, van Garderen-Hoetmer A, Wijnands MV.
Dietary fat and carcinogenesis. Mutat Res 1999;443:111-27.
5. Nason FG, Nelson BE. Estrogen and progesterone in breast and
gynecologic cancers. Etiology, therapeutic role, and hormone replacement.
Obstet Gyn Clin N Am 1994;21:245-70.
6. Herrinton LJ, Stanford JL, Schwartz SM, Weiss NS. Ovarian cancer
incidence among Asian migrants to the United States and their descendants.
J Natl Cancer Inst 1994.;86:1336-9.
7. Persson I. Estrogens in the causation of breast, endometrial
and ovarian cancers - evidence and hypotheses from epidemiological findings.
J Steroid Biochem Mol Biol 2000;74:357-64.
8. Adami HO, Hunter D, Trichopoulos D. Textbook of Cancer Epidemiology;
New York: Oxford University Press, 2002. (302p for breast, 360p corpus
uteri, and 380p for ovarian cancer).
9. Key TJ. Hormones and cancer in human. Mutat Res 1995;333:59-67.
10. Armstrong B, Doll R. Environmental factors and cancer incidence
and mortality in different countries, with special reference to dietary
practices. Int J Cancer 1975;15:617-31.
11. Parkin DM, Whelan SL, Ferlay J, Teppo L and Thomas DB, eds.
Cancer Incidence in Five Continents. Vol. VIII. IARC Sci Publ 2002;155,
Lyon, IARC.
12. FAOSTAT Database Collections. http://apps.fao.org/cgi-bin/nph-db.
pl?subset=nutrition/
13. Ferlay J, Bray F, Pisani P, and Parkin DM. GLOBOCAN 2000: Cancer
Incidence, Mortality and Prevalence Worldwide. Lyon: IARC, 2001.
14. Rose DP, Boyar AP, Wynder EL. International comparisons of
mortality rates for cancer of the breast, ovary, prostate, and colon,
and per capita food consumption. Cancer 1986;58:2363-71.
15. Jin F, Shu XO, Devesa SS, Zheng V, Blot WJ, Gao YT. Incidence
trends for cancer of the breast, ovary, and corpus uterin in urban Shanghai,
1972-1989, Cancer Causes Control 1993;4:355-60.
16. Nagata C, Kawakami N, Shimizu H. Trends in the incidence rate
and risk factors for breast cancer in Japan. Breast Cancer Res Treat 1997;44:75-82.
17. Talamini R, La Vecchia C, Decarli A, et al. Social factors,
diet and breast cancer in a northern Italian population. Br J Cancer 1984;49:723-9.
18. La Vecchia C, Pampallona S. Age at first birth, dietary practices
and breast cancer mortality in various Italian regions. Oncology 1986.;43:1-6.
19. Boyd NF, Martin LJ, Noffel M, Lockwood GA, Trichler DL. A meta-analysis
of studies of dietary fat and breast cancer risk. Br J Cancer 1993;68:627-36.
20. Levi F, La Vecchia C, Gulie C, Negri E. Dietary factors and
breast cancer risk in Vaud, Switzerland. Nutr Cancer 1993;19:327-35.
21. Gaard M, Tretli S and L�ken EB. Dietary fat and the risk of
breast cancer: a prospective study of 25,892 Norwegian women. Int J Cancer
1995;63:13-17.
22. M�nnist� S, Pietinen P, Virtanen M, Kataja, Uusitupa M. Diet
and the risk of breast cancer in a case-control study: does the threat
of disease have an influence on recall bias? J Clin Epidemiol 1999;52:429-39.
23. Willett WC. Diet and breast cancer. J Int Med 2001;249:395-411.
24. Mettlin CJ, Schoenfeld ER, Natarajan N. Patterns of milk consumption
and risk of cancer. Nutr Cancer 1990;13:89-99.
25. Leung GM, Thach TQ, Lam TH, et al. Trends in breast cancer
incidence in Hong Kong between 1973 and 1999: an age-period-cohort analysis.
Br J Cancer 2002;87:982-8.
26. Toniolo PG, Levitz M, Zeleniuch-Jacquotte A, et al. A prospective
study of endogenous estrogens and breast cancer in postmenopausal women.
J Natl Cancer Inst 1995;87:190-7.
27. Hartmann S, Lacorn M, Steinhart H. Natural occurrence of steroid
hormones in food. Food Chem 1998;62:7-20.
28. Ganmaa D, Wang PY, Qin LQ, Hoshi K, Sato A. Is milk responsible
for male reproductive disorders? Med Hypoth 2001;57:510-4.
29. Roberts KD, Rochefort JG, Bleau G, Chapdelaine A. Plasma estrone
sulfate levels in postmenopausal women. Steroids 1980;35:179-87.
30. Moore AB, Bottoms GD, Coppoc GL, Pohland RC, Roesel OF. Metabolism
of estrogens in the gastrointestinal tract of swine. I. Instilled estradiol.
J Animal Sci 1982;55:124-34.
31. Jasonni VM, Naldi S, Ciotti P, Bulletti C, Flamigni C. Comparative
metabolism of oestrone sulphate after oral and intravenous administration
in post-menopausal women. Maturitas. 1987;9:201-5.
32. Remesar X, Tang V, Ferrer E, et al. Estrone in food: a factor
influencing the development of obesity? Eur J Nutr 1999;38:247-53.
33. Qin LQ, Xu JY, Wang PY, et al. Low-Fat Milk Promotes the Development
of 7,12-Dimethylbenz(a)anthracene (DMBA)-Induced Mammary Tumors in Rats.
Int J Cancer 2004;110:491-6.
34. Hill P, Wynder EL, Garnes H, Walker AR. Environmental factors,
hormone status, and prostatic cancer. Prev Med 1980;9:657-66.
35. Bernstein L, Ross RK. Endogenous hormones and breast cancer
risk. Epidemiol Rev 1993;15:48-65.
36. Outwater JL, Nicholson A, Barnard N. Dairy products and breast
cancer: the IGF-I, estrogen, and bGH hypothesis. Med Hypoth 1997;48:453-61.
37. Holly JM, Gunnell DJ, Davey Smith G. Growth hormone, IGF-I
and cancer. Less intervention to avoid cancer? More intervention to prevent
cancer? J Endocrinol 1999;162:321-30.
38. Macaulay VM. Insulin-like growth factors and cancer. Br J Cancer
1992;65:311-20
39. Heaney RP, McCarron DA, Dawson-Hughes B, et al. Dietary changes
favorably affect bone remodeling in older adults. J Am Diet Ass 1999;99:1228-33.
40. Key TJ, Allen NE, Verkasalo PK, Banks E. Energy balance and
cancer: the role of sex hormones. Proc Nutr Soc 2001;60:81-9.
41. Dal Maso L, Franceschi S, Negri E, et al. Body size indices
at different ages and epithelial ovarian cancer risk. Eur J Cancer 2002;38:1769-74.
42. MacDonald PC, Edman CD, Hemsell DL, Porter JC, Siiteri PK.
Effect of obesity on conversion of plasma androstenedione to estrone in
postmenopausal women with and without endometrial cancer. Am J Obst Gynecol
1978;130:448-55.
43. Rossouw JE, Anderson GL, Prentice RL, et al. Writing Group
for the Women's Health Initiative Investigators. Risks and benefits of
estrogen plus progestin in healthy postmenopausal women: principal results
From the Women's Health Initiative randomized controlled trial. JAMA 2002;288:321-33.
44. Beral V, Million Women Study Collaborators. Breast cancer and
hormone-replacement therapy in the Million Women Study. Lancet 2003;362:419-27.
45. Kushi LH, Mink PJ, Folsom AR, et al. Prospective study of diet
and ovarian cancer. Am J Epidemiol 1999;149:21-31.
46. Cramer DW. Lactase persistence and milk consumption as determinants
of ovarian cancer risk. Am J Epidemiol 1989;130:904-10.
47. Kaufman FR, Kogut MD, Donnell GN, Goebelsmann U, March C, Koch
R. Hypergonadotropic hypogonadism in female patients with galactosemia.
N Engl J Med 1981;304(17):994-8.
48. Cramer DW, Harlow BL, Willett WC, et al. Galactose consumption
and metabolism in relation to the risk of ovarian cancer. Lancet 1989;2(8654):66-71.
49. Risch HA, Jain M, Marrett LD, Howe GR. Dietary fat intake and
risk of epithelial ovarian cancer. J Natl Cancer Inst 1994;86:1409-15.
50. Cramer DW, Greenberg ER, Titus-Ernstoff L, et al. A case-control
study of galactose consumption and metabolism in relation to ovarian cancer.
Cancer Epidemiology, Biomarkers Prev 2000;9:95-101.
51. Larsson SC, Bergkvist L, Wolk A. Milk and lactose intakes and
ovarian cancer risk in the Swedish Mammography Cohort. Am J Clin Nutr
2004;80:1353-7.
52. Cramer DW, Welch WR, Hutchison GB, Willett W, Scully RE. Dietary
animal fat in relation to ovarian cancer risk. Obstet Gynecol 1984;63:833-8.
53. La Vecchia C, Decarli A, Negri E, et al. Dietary factors and
the risk of epithelial ovarian cancer. J Natl Cancer Inst 1987;79:663-9.
54. Mettlin CJ, Piver MS. A case-control study of milk-drinking
and ovarian cancer risk. Am J Epidemiol 1990;132:871-6.
55. Webb PM, Bain CJ, Purdie DM, Harvey PW, Green A. Milk consumption,
galactose metabolism and ovarian cancer (Australia). Cancer Causes &
Control 1998;9:637-44.
56. Karasik A, Menczer J, Pariente C, Kanety H. Insulin-like growth
factor-I (IGF-I) and IGF-binding protein-2 are increased in cyst fluids
of epithelial ovarian cancer. J Clin Endocr Metab 1994;78:271-6.
57. Flyvbjerg A. Mogensen O. Mogensen B. Nielsen OS. Elevated serum
insulin-like growth factor-binding protein 2 (IGFBP-2) and decreased IGFBP-3
in epithelial ovarian cancer: correlation with cancer antigen 125 and
tumor-associated trypsin inhibitor. J Clin Endocr Metab 1997;82:2308-13.
58. Key TJ, Pike MC. The dose-effect relationship between 'unopposed'
oestrogens and endometrial mitotic rate: its central role in explaining
and predicting endometrial cancer risk. Br J Cancer 1988;57:205-12.
59. Akhmedkhanov A, Zeleniuch-Jacquotte A, Toniolo P. Role of exogenous
and endogenous hormones in endometrial cancer: review of the evidence
and research perspectives. Ann NY Acad Sci 2001;943:296-315.
60. Liehr JG. Genotoxicity of the steroidal oestrogens oestrone
and oestradiol: possible mechanism of uterine and mammary cancer development.
Hum Reprod Update 2001;7:273-81.
61. Kleinman D, Karas M, Danilenko M, et al. Stimulation of endometrial
cancer cell growth by tamoxifen is associated with increased insulin-like
growth factor (IGF)-I induced tyrosine phosphorylation and reduction in
IGF binding proteins. Endocrinology 1996;137:1089-95.
62. Ayabe T, Tsutsumi O, Sakai H, et al. Increased circulating
levels of insulin-like growth factor-I and decreased circulating levels
of insulin-like growth factor binding protein-1 in postmenopausal women
with endometrial cancer. Endocr J 1997;44:419-24.
63. Reynolds RK, Hu C, Baker VV. Transforming growth factor-alpha
and insulin-like growth factor-I, but not epidermal growth factor, elicit
autocrine stimulation of mitogenesis in endometrial cancer cell lines.
Gynecol Oncol 1998;70:202-9.